
terça-feira, 27 de março de 2012


Gametogênese:
Processo que ocorre nas gônadas animais, formando os gametas e q subdivide-se e dois grupos distintos:
Formação dos gametas masculinos, originando os espermatozóides;
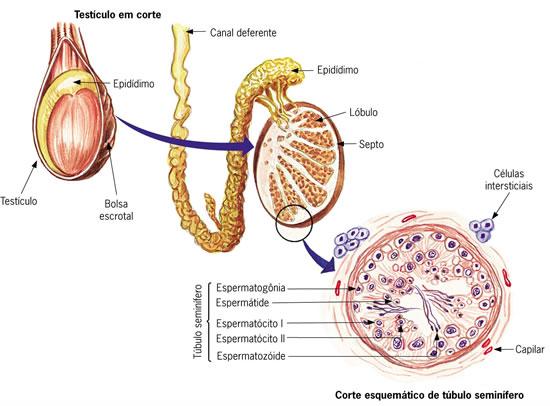
Fase de Multiplicação: Nos testículos, localizam-se os tubos seminíferos, onde se encontram as espermatogônias (2n), onde as mesmas se multiplicam por mitose. Após a multiplicação, ocorre a fase de crescimento, onde algumas espermatogônias crescem e duplicam seus cromossomos, tornando-se espermatócitos I, onde os mesmos sofrem meiose, originando células haplóides denominadas espermatócitos II, que também sofrerá meiose, formando células haplóides chamadas de espermátides. As duplicações meióticas sofridas pelos espermatócitos são denominadas de Fase de Maturação.
Após a fase de maturação, inicia-se a Especiação ou Diferenciação, onde as espermátides perdem praticamente todo seu citoplasma, e começam a desenvolver o flagelo, a partir do centríolo. Formado o flagelo, apresenta-se uma nova estrutura: o espermatozóide.
Na cabeça do espermatozóide, encontra-se o acrossomo, q é formado pelo Complexo de Golgi, contendo enzimas que facilitarão a penetração do gameta no óvulo. Além do acrossomo, encontra-se também o núcleo, onde se armazena os cromossomos paternos.
Já no inicio do flagelo, encontram-se as mitocôndrias, para o fornecimento de energia para o espermatozóide.
 Oogênese:
Oogênese:
formação dos gametas femininos, originando os óvulos.
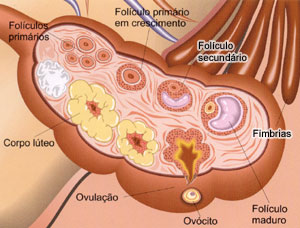
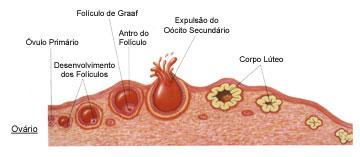
Fase de Multiplicação: Ainda na vida intra-uterina, células germinativas primordiais migram para o ovário e sofrem diferenciação, formando oogônias (2n).
Após todas as divisões, cada oogônia passa por um período de crescimento, síntese e acúmulo de materiais de reserva. A partir daí, as oogônias passam a denominar-se de oócitos I ou oócitos de primeira ordem, sendo encontrados nos folículos primários. Esse desenvolvimento é conhecido como Fase de Crescimento.
Já na Fase de Maturação, ocorrem mais divisões por meiose formando os gametas, onde na primeira divisão formam-se duas células, o oócito II ou oócio de segunda ordem e o primeiro glóbulo polar. Na segunda divisão meiótica, o oócito II forma mais duas células, o óvulo e o segundo glóbulo polar, este que, se degenera junto com o primeiro glóbulo polar.
MOREIRA,Catarina. Oogênese. Disponível em :http://wikiciencias.casadasciencias.org/index.php/Oog%C3%A9nese. Acessado em 27 de março de 2012.
BOS- MIKICH, Adriana. Oogênese. Disponível em:http://www.ufrgs.br/gametaembriao/oogenese.htm. Acessado em 27 de março de 2012
Biologia Geral- Henning&Ferraz
Meiose
O que é meiose?
A meiose é um processo de divisão celular em que uma célula diplóide (2n) forma quatro células haplóides (n). A meiose consiste em duas divisões celulares, acompanhadas por uma só duplicação cromossômica.

Organismos simples podem reproduzir-se através de divisões simples. Este tipo de reprodução assexuada é simples e direta e produz organismos geneticamente iguais. A reprodução sexual por sua vez, envolve uma mistura de genomas de 2 indivíduos, para produzir um indivíduo que diferem geneticamente de seus parentais.
O ciclo reprodutivo sexual envolve a alternância de gerações de células haplóides, com gerações de células diplóides. A mistura de genomas é realizada pela fusão de células haplóides que formam células diplóides. Posteriormente novas células diplóides são geradas quando os descendentes de células diplóides se dividem pelo processo de meiose.
Com exceção dos cromossomos que determinam o sexo, um núcleo de célula diplóide contém 2 versões similares de cada cromossomo autossomo, um cromossomo paterno e 1 cromossoma materno. Essas duas versões são chamadas de homologas, e na maioria das células possuem existência como cromossomos independentes. Essas duas versões são denominadas de homólogos. Quando o DNA é duplicado pelo processo de replicação, cada um desses cromossomos é replicado dando origem as cromátides que são então separadas durante a anáfase e migram para os polos celulares. Desta maneira cada célula filha recebe uma cópia do cromossomo paterno e uma cópia do cromossoma materno.
Vimos que a mitose resulta em células com o mesmo número de cromossomas, se ocorre - se a fusão dessas células, teríamos como resultado células com o dobro de cromossomas e isso ocorreria em progressão. Exemplificando: O homem possui 46 cromossomas, a fusão resultaria em uma célula com 92 cromossomas. A meiose desenvolveu-se para evitar essa progressão.
A meiose ( meioum = diminuir ) ocorre nas células produtoras de gametas. Os gametas masculinos e femininos ( espermatozóides e óvulos ) que são produzidos nos testículos e ovários respectivamente as gônadas femininas e masculinas. Os gametas se originam de células denominadas espermatogonias e ovogonias.
A meiose é precedida por um período de intérfase ( G1, S, G2 ) com eventos semelhantes aos observados na mitose.
As espermatogônias e ovogônias, que são células diplóides, sofrem sucessivas divisões mitóticas. As células filhas dessas células desenvolvem ciclo celular, e num determinado momento da fase G2 do ciclo celular ocorrem alterações que levam as células a entrar em meiose e darem origem a células háploides ou seja células que possuem a metade do número (n) de cromossomas da espécie. A regulação do processo meiótico inicia-se durante a fase mitótica, onde observam-se: 1) Período S longo; 2) aumento do volume nuclear. Experimentalmente demonstra-se que eventos decisivos ocorrem em G2, devido a ativação de sítios únicos para a meiose. Podemos definir meiose como sendo o processo pelo qual número de cromossomos é reduzido a metade.
Na meiose o cromossomo produzido possui apenas a metade do número de cromossomos, ou seja somente um cromossomo no lugar de um par de homólogos. O gameta é dotado de uma cópia do cromossoma materno ou paterno.
A meiose é um processo que envolve 2 divisões celulares com somente uma duplicação de cromossomas.
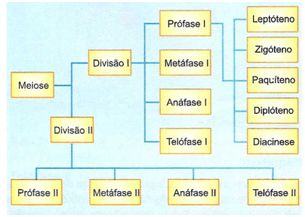
Fases da Meiose
A meiose ocorre apenas nas células das linhagens germinativas masculina e feminina e é constituída por duas divisões celulares: Meiose I e Meiose II.
O esquema geral da meiose
A meiose envolve duas divisões celulares. A primeira divisão meiótica é chamada reducional, pois reduz o número de cromossomos de um estado diplóide (2n) para o haplóide (n). A segunda divisão é chamada equacional e mantém o número haplóide. O processo geral obedece ao seguinte esquema:
Meiose divisão I - Divisão reducional
A meiose I é subdividida em quatro fases, denominadas: Prófase I, Metáfase I, Anáfase I, Telófase I
PRÓFASE I
A prófase I é de longa duração e muito complexa. Os cromossomos homólogos se associam formando pares, ocorrendo permuta (crossing-over) de material genético entre eles. Vários estágios são definidos durante esta fase: Leptóteno, Zigóteno, Paquíteno, Diplóteno e Diacinese.

O crossing-over
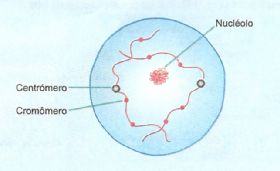
Leptóteno
Os cromossomos tornam-se visíveis como delgados fios que começam a se condensar, mas ainda formam um denso emaranhado. Nesta fase inicial , as duas cromátides- irmãs de cada cromossomo estão alinhadas tão intimamente que não são ditinguíveis.
Zigóteno
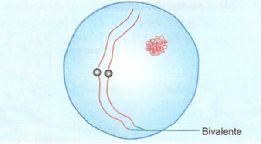
Os cromossomos homólogos começam a combinar-se estreitamente ao longo de toda a sua extensão. O processo de pareamento ou sinapse é muito preciso.
Paquíteno
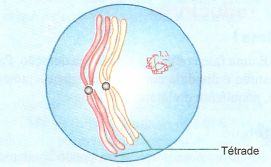
Os cromossomos tornam-se bem mais espiralados. O pareamento é completo e cada par de homólogos aparece como um bivalente ( às vezes denominados tétrade porque contém quatro cromátides)
Neste estágio ocorre o crossing-over, ou seja, a troca de segmentos homólogos entre cromátides não irmãs de um par de cromossomos homólogos.
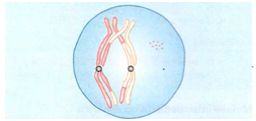
Diplóteno
Ocorre o afastamento dos cromossomos homólogos que constituem os bivalentes. Embora os cromossomos homólogos se separem, seus centrômeros permanecem intactos, de modo que cada conjunto de cromátides-irmãs continua ligado inicialmente. Depois, os dois homólogos de cada bivalente mantêm-se unidos apenas nos pontos denominados quiasmas (cruzes).
.jpg)
Diacinese
Neste estágio os cromossomos atingem a condensação máxima.
METÁFASE I
Há o desaparecimento da membrana nuclear. Forma-se um fuso e os cromosomos pareados se alinham no plano equatorial da célula com seus centrômeros orientados para pólos diferentes.
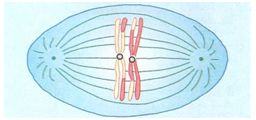
ANÁFASE I
Os dois membros de cada bivalente se separam e seus respectivos centrômeros com as cromátides-irmãs fixadas são puxados para pólos opostos da célula.
Os bivalentes distribuem-se independentemente uns dos outros e, em consequência, os conjuntos paterno e materno originais são separados em combinações aleatórias.
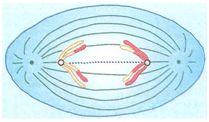
TELÓFASE I
Nesta fase os dois conjuntos haplóides de cromossomos se agrupam nos pólos opostos da célula.
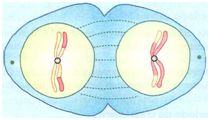
Meiose divisão II - Divisão equacional
A meiose II tem início nas células resultantes da telófase I, sem que ocorra a Intérfase. A meiose II também é constituída por quatro fases:
PRÓFASE II
É bem simplificada, visto que os cromossomos não perdem a sua condensação durante a telófase I. Assim, depois da formação do fuso e do desaparecimento da membrana nuclear, as células resultantes entram logo na metáfase II.
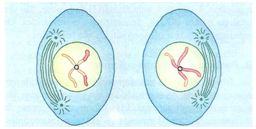
METÁFASE II
Os 23 cromossomos subdivididos em duas cromátides unidas por um centrômero prendem-se ao fuso.
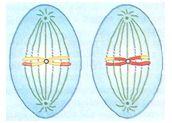
ANÁFASE II
Após a divisão dos centrômeros as cromátides de cada cromossomo migram para pólos opostos.
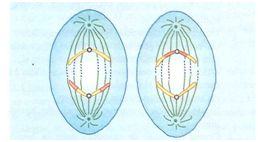
TELÓFASE II
Forma-se uma membrana nuclear ao redor de cada conjunto de cromátides.
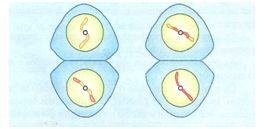
DUARTE, Thais. Mitose e Meiose. Disponível em : http://www.coladaweb.com/biologia/biologia-celular/mitose-e-meiose. Acessado em 27 de março de 2012.
Divisões celulares
Na mitose, a divisão de uma “célula-mãe” duas “células-filhas” geneticamente idênticas e com o mesmo número cromossômico que existia na célula-mãe. Uma célula n produz duas células n, uma célula 2n produz duas células 2n etc. Trata-se de uma divisão equacional.
Já na meiose, a divisão de uma “célula-mãe” 2n gera “células-filhas” n, geneticamente diferentes. Neste caso, como uma célula 2n produz quatro células n, a divisão é chamada reducional.
A interfase – A fase que precede a mitose
Interfase é o período que precede qualquer divisão celular, sendo de intensa atividade metabólica. Nesse período, há a preparação para a divisão celular, que envolve a duplicação da cromatina, material responsável pelo controle da atividade da célula. Todas as informações existentes ao longo da molécula de DNA são passadas para a cópia, como se correspondessem a uma cópia fotográfica da molécula original.
Em pouco tempo, cada célula formada da divisão receberá uma cópia exata de cada cromossomo da célula se dividiu.
As duas cópias de cada cromossomo permanecem juntas por certo tempo, unidas pelo centrômero comum, constituindo duas cromátides de um mesmo cromossomo. Na interfase, os centríolos também se duplicam.
A interfase e a Duplicação do DNA
Houve época em que se falava que a interfase era o período de “repouso” da célula. Hoje, sabemos, que na realidade a interfase é um período de intensa atividade metabólica no ciclo celular: é nela que se dá a duplicação do DNA, crescimento e síntese. Costuma-se dividir a interfase em três períodos distintos: G1, S e G2.
O intervalo de tempo em que ocorre a duplicação do DNA foi denominado de S (síntese) e o período que antecede é conhecido como G1 (G1 provém do inglês gap, que significa “intervalo”). O período que sucede o S é conhecido como G2.

O ciclo celular todo, incluindo a interfase (G1, S, G2) e a mitose (M) – prófase, metáfase, anáfase e telófase – pode ser representado em um gráfico no qual se coloca a a quantidade da DNA na ordenada (y) e o tempo na abscissa (x). Vamos supor que a célula que vai se dividir tenha, no período G1, uma quantidade 2C de DNA (C é uma unidade arbitrária). O gráfico da variação de DNA, então, seria semelhante ao da figura abaixo.

Nas células, existe uma espécie de “manual de verificação de erros” que é utilizado em algumas etapas do ciclo celular e que é relacionado aos pontos de checagem. Em cada ponto de checagem a célula avalia se é possível avançar ou se é necessário fazer algum ajuste, antes de atingir a fase seguinte. Muitas vezes, a escolha é simplesmente cancelar o processo ou até mesmo conduzir a célula à morte
As fases da mitose
A mitose é um processo contínuo de divisão celular, mas, por motivos didáticos, para melhor compreendê-la, vamos dividi-la em fases: prófase, metáfase, anáfase e telófase Alguns autores costumam citar uma quinta fase – a prometáfase – intermediária entre a prófase emetáfase. O final da mitose, com a separação do citoplasma, é chamado de citocinese.
Prófase – Fase de início (pro = antes)

- Os cromossomos começam a ficar visíveis devido à espiralação.
- O nucléolo começa a desaparecer.
- Organiza-se em torno do núcleo um conjunto de fibras (nada mais são do que microtúbulos) originadas a partir dos centrossomos, constituindo o chamado fuso de divisão (ou fuso mitótico).
- Embora os centríolos participem da divisão, não é deles que se originam as fibras do fuso. Na mitose em célula animal, as fibras que se situam ao redor de cada par de centríolos opostas ao fuso constituem o áster (do grego,aster = estrela).
- O núcleo absorve água, aumenta de volume e a carioteca se desorganiza.
- No final da prófase, curtas fibras do fuso, provenientes do centrossomos, unem-se aos centrômeros. Cada uma das cromátides-irmãs fica ligada a um dos pólos da célula.
Note que os centrossomos ainda estão alinhados na região equatorial da célula, o que faz alguns autores designarem essa fase de prometáfase. - O núcleo absorve água, aumenta de volume e a carioteca se desorganiza.
Metáfase – Fase do meio (meta = no meio)
- Os cromossomos atingem o máximo em espiralação, encurtam e se localizam na região equatorial da célula.
- No finalzinho da metáfase e início da anáfase ocorre a duplicação dos centrômeros.

Anáfase – Fase do deslocamento (ana indica movimento ao contrário)
- As fibras do fuso começam a encurtar. Em conseqüência, cada lote de cromossomos-irmãos é puxado para os pólos opostos da célula.
Como cada cromátide passa a ser um novo cromossomo, pode-se considerar que a célula fica temporariamente tetraplóide.

Telófase – Fase do Fim (telos = fim)
- Os cromossomos iniciam o processo de desespirilação.
- Os nucléolos reaparecem nos novos núcleos celulares.
- A carioteca se reorganiza em cada núcleo-filho.
- Cada dupla de centríolos já se encontra no local definitivo nas futuras células-filhas.

Citocinese – Separando as células
A partição em duas copias é chamada de citocinese e ocorre, na célula animal, de fora para dentro, isto é, como se a célula fosse estrangulada e partida em duas (citocinese centrípeta). Há uma distribuição de organelas pelas duas células-irmãs. Perceba que a citocinese é, na verdade a divisão do citoplasma. Essa divisão pode ter início já na anáfase, dependendo da célula.

A mitose é um tipo de divisão muito freqüente entre os organismos da Terra atual. Nos unicelulares, serve à reprodução assexuada e à multiplicação dos organismos. Nos pluricelulares, ela repara tecidos lesados, repões células que normalmente morrem e também está envolvida no crescimento.
No homem, a pele, a medula óssea e o revestimento intestinal são locais onde a mitose é freqüente. Nem todas as células do homem, porém, são capazes de realizar mitose. Neurônios e célula musculares são dois tipos celulares altamente especializados em que não ocorre esse tipo de divisão (ocorre apenas na fase embrionária). Nos vegetais, a mitose ocorre em locais onde existem tecidos responsáveis pelo crescimento, por exemplo, na ponta de raízes, na ponta de caules e nas gemas laterais.

Divisões celulares. Disponível em: <http://www.sobiologia.com.br/conteudos/Citologia 2/nucleo6.php>. Acesso em :27 mar. 2012.
domingo, 25 de março de 2012
A fronteira das células
No mundo de hoje, é comum pensarmos em um país como sendo uma porção de terra delimitada espacialmente das demais pela presença de uma fronteira. Vamos pensar no caso do Brasil. Estamos rodeados de mar em metade do nosso território e, na outra metade, fazemos fronteira terrestre com outros nove países da America do Sul. Em suas fronteiras, todos os países instalam uma alfândega, que é uma repartição governamental de controle do movimento de entradas e saídas das pessoas e de mercadorias para o exterior ou deles provenientes.
Com as células não é diferente. Cada uma delas tem uma “área de fronteira”, representada pela membrana plasmática e, nesta área, as células também possuem o seu “posto alfandegário”, as proteínas. Assim como nas aduanas das fronteiras entre os países, essas proteínas são as responsáveis pelo reconhecimento de substâncias vindas de dentro ou de fora da célula como, por exemplo, hormônios.
O trabalho realizado por uma célula é semelhante ao que acontece em uma fábrica, como a de televisores, por exemplo. Através de portões, dá-se a entrada de diversos tipos de peças destinadas as linhas de montagem. Para a fabricação e a montagem dos aparelhos, são necessários energia e operários habilitados. É preciso, ainda, um setor de embalagem para preparar a expedição do que é produzido e uma diretoria para comandar todo o complexo fabril e manter o relacionamento com o mundo externo. Tudo dentro dos limites representados pelo muro da fábrica.

ORGANELAS
Uma organela pode ser definida como uma determinada parte do citoplasma responsável por uma ou mais definições especiais

Membrana celular
(ou membrana plasmática ou membrana citoplasmática ou plasmalema)
Toda a célula, seja procarionte ou eucarionte, apresenta uma membrana que isola do meio exterior: a membrana plasmática. A membrana plasmática é tão fina (entre 6 a 9 nm) que os mais aperfeiçoados microscópios ópticos não conseguiram torná-la visível. Foi somente após o desenvolvimento da microscopia eletrônica que a membrana plasmática pode ser observada. Nas grandes ampliações obtidas pelo microscópio eletrônico, cortes transversais da membrana aparecem como uma linha mais clara entre duas mais escuras, delimitando o contorno de cada célula.
Constituição química da membrana plasmática
Estudos com membranas plasmáticas isoladas revelam que seus componentes mais abundantes são fosfolipídios, colesterol e proteínas. É por isso que se costumam dizer que as membranas plasmáticas têm constituição lipoprotéica.

Uma vez identificados os fosfolipídios e as proteínas como os principais componentes moleculares da membrana, os cientistas passaram a investigar como estas substâncias estavam organizadas.
O modelo do mosaico fluído
A disposição das moléculas na membrana plasmática foi elucidada recentemente, sendo que os lipídios formam uma camada dupla e contínua, no meio da qual se encaixam moléculas de proteína. A dupla camada de fosfolipídios é fluida, de consistência oleosa, e as proteínas mudam de posição continuamente, como se fossem peças de um mosaico. Esse modelo foi sugerido por dois pesquisadores, Singer e Nicholson, e recebeu o nome de Modelo Mosaico Fluido.
Os fosfolipídios têm a função de manter a estrutura da membrana e as proteínas têm diversas funções. As membranas plasmáticas de um eucariócitos contêm quantidades particularmente grande de colesterol. As moléculas de colesterol aumentam as propriedades da barreira da bicamada lipídica e devido a seus rígidos anéis planos de esteróides diminuem a mobilidade e torna a bicamada lipídica menos fluida.

Funções das proteínas na membrana plasmática
As proteínas da membrana plasmática exercem grandes variedades de funções: atuam preferencialmente nos mecanismos de transporte, organizando verdadeiros túneis que permitem a passagem de substâncias para dentro e para fora da célula, funcionam como receptores de membrana, encarregadas de receber sinais de substâncias que levam alguma mensagem para a célula, favorecem a adesão de células adjacentes em um tecido, servem como ponto de ancoragem para o citoesqueleto.
- Proteínas de adesão: em células adjacentes, as proteínas da membrana podem aderir umas às outras.
- Proteínas que facilitam o transporte de substâncias entre células.
- Proteínas de reconhecimento: determinadas glicoproteínas atuam na membrana como um verdadeiro “selo marcador”, sendo identificadas especificamente por outras células.
- Proteínas receptoras de membrana.
- Proteínas de transporte: podem desempenhar papel na difusão facilitada, formando um canal por onde passam algumas substâncias, ou no transporte ativo, em que há gasto de energia fornecida pela substância ATP. O ATP (adenosina trifosfato) é uma molécula derivada de nucleotídeo que armazena a energia liberada nos processos bioenergéticos que ocorrem nas células (respiração aeróbia, por exemplo). Toda vez que é necessária energia para a realização de uma atividade celular (transporte ativo, por exemplo) ela é fornecida por moléculas de ATP.
- Proteínas de ação enzimática: uma ou mais proteínas podem atuar isoladamente como enzima na membrana ou em conjunto, como se fossem parte de uma “linha de montagem” de uma determinada via metabólica.
- Proteínas com função de ancoragem para o citoesqueleto.
Núcleo

Normalmente apresentam forma ovoide ou esférica, com diâmetro médio
igual a 5μm, porém também manifestando morfologia lobular: bilobulados ou
multilobulados, observados em células de defesa (alguns tipos de leucócitos).
Entretanto, existem células anucleadas, por exemplo, as hemácias dos seres humanos. Desta forma, de acordo com a diferenciação entre os tecidos, células diferentes em um mesmo organismo podem variar quanto ao número de núcleos, sendo: mononucleadas, possuindo somente um núcleo (células epiteliais); binucleadas, com dois núcleos (células hepáticas), e multinucleadas, contendo vários núcleos (células musculares).
Características e função da membrana nuclear da célula:
- Formada por duas camadas lipoproteicas;
- Apresenta numerosos poros comunicantes com o hialoplasma, por onde saem e entram substâncias moleculares;
- Proteção do material genético;
- E barreira física que limita a região reguladora do metabolismo, através do processo de transcrição.
Entretanto, existem células anucleadas, por exemplo, as hemácias dos seres humanos. Desta forma, de acordo com a diferenciação entre os tecidos, células diferentes em um mesmo organismo podem variar quanto ao número de núcleos, sendo: mononucleadas, possuindo somente um núcleo (células epiteliais); binucleadas, com dois núcleos (células hepáticas), e multinucleadas, contendo vários núcleos (células musculares).
Características e função da membrana nuclear da célula:
- Formada por duas camadas lipoproteicas;
- Apresenta numerosos poros comunicantes com o hialoplasma, por onde saem e entram substâncias moleculares;
- Proteção do material genético;
- E barreira física que limita a região reguladora do metabolismo, através do processo de transcrição.
Contudo, mesmo uma célula nucleada, dependendo do estágio de seu ciclo celular, pode admitir distintos comportamentos: durante a interfase, período de síntese intensa, o núcleo apresenta aspecto evidente, enquanto no período de multiplicação (divisão - mitose ou meiose) tanto a carioteca quanto o nucléolo se desintegram, reaparecendo no final deste evento.

Mitocôndrias
Estrutura e função das mitocôndria
As mitocôndrias estão imersas no citosol, entre as diversas bolsas e filamentos que preenchem o citoplasma das células eucariontes. Elas são verdadeiras “casas de força” das células, pois produzem energia para todas as atividades celulares.
As mitocôndrias foram descobertas em meados do século XIX, e, durante décadas, sua existência foi questionada por alguns citologistas. Somente em 1890 foi demonstrada, de modo incontestável, a presença de mitocôndrias no citoplasma celular. O termo “mitocôndria” (do grego, mitos, fio, e condros, cartilagem) surgiu em 1898, possivelmente como referência ao aspecto filamentoso e homogêneo (cartilaginoso) dessas organelas em alguns tipos de células, quando observadas ao microscópio óptico.
As mitocôndrias, cujo número varia de dezenas até centenas, dependendo do tipo de célula, estão presentes praticamente em todos os seres eucariontes, sejam animais, plantas, algas, fungos ou protozoários.
Estrutura interna das mitocôndrias
As mitocôndrias são delimitadas por duas membranas lipoprotéicas semelhantes às demais membranas celulares. Enquanto a membrana externa é lisa, a membrana interna possui inúmeras pregas – as cristas mitocondriais – que se projetam para o interior da organela.

Estrutura interna das mitocôndrias
As mitocôndrias são delimitadas por duas membranas lipoprotéicas semelhantes às demais membranas celulares. Enquanto a membrana externa é lisa, a membrana interna possui inúmeras pregas – as cristas mitocondriais – que se projetam para o interior da organela.
A cavidade interna das mitocôndrias é preenchida por um fluido denominado matriz mitocondrial, onde estão presentes diversas enzimas, além de DNA e RNA e pequenos ribossomos e substâncias necessárias à fabricação de determinadas proteínas.
A respiração celular
No interior das mitocôndrias ocorre a respiração celular, processo em que moléculas orgânicas de alimento reagem com gás oxigênio (O2), transformando-se em gás carbônico (CO2) e água (H2O) e liberando energia.
C6H12O6 + O2 -> 6 CO2 + 6 H2O + energia
|
A energia liberada na respiração celular é armazenada em uma substância chamada ATP (adenosina trifosfato), que se difunde para todas as regiões da célula, fornecendo energia para as mais diversas atividades celulares. O processo de respiração celular será melhor explicado na seção de Metabolismo energético.
A origem das mitocôndrias
Toda mitocôndria surge da reprodução de uma outra mitocôndria. Quando a célula vai se dividir, suas mitocôndrias se separam em dois grupos mais ou menos equivalentes, que se posicionam em cada um dos lados do citoplasma.

Ao final da divisão cada um dos grupos fica em uma célula-filha. Posteriormente, no decorrer do crescimento das células, as mitocôndrias se duplicam e crescem, restabelecendo o número original.
As mitocôndrias do espermatozóide penetram no óvulo durante a fecundação e degeneram-se logo em seguida, portanto, as mitocôndrias presentes na célula-ovo são originárias exclusivamente da mãe. As mitocôndrias ovulares, que se multiplicam sempre que a célula se reproduz, são as ancestrais de todas, as mitocôndrias presentes em nossas células.
Muitos cientistas estão convencidos de que as mitocôndrias são descendentes de seres procariontes primitivos que um dia se instalaram no citoplasma das primeiras células eucariontes. Existem evidências que apóiam essa hipótese, tais como o fato de as mitocôndrias apresentarem material genético mais parecido com a das bactérias do que com a das células eucariontes em que se encontram

O mesmo ocorre com relação a maquinaria para a síntese de proteínas: os ribossomos mitocondriais são muito semelhantes aos das bactérias e bem diferentes dos ribossomos presentes no citoplasma das células eucariontes.

Aparelho de Golgi
A denominação aparelho ou complexo de Golgi é uma homenagem ao citologista italiano Camilo Golgi, que, em 1898, descobriu essa estrutura citoplasmática. Ao verificar que certas regiões com citoplasma celular se coravam por sais de ósmio de prata, Golgi imaginou que ali deveria existir algum tipo de estrutura, posteriormente confirmada pela microscopia eletrônica.
Dictiossomos
O aparelho de Golgi está presente em praticamente todas as células eucariontes, e consiste de bolsas membranosas achatadas, empilhadas como pratos. Cada uma dessas pilhas recebe o nome dedictiossomo. Nas células animais, os dictiossomos geralmente se encontram reunidos em um único local, próximo ao núcleo. Nas células vegetais, geralmente há vários dictiossomos espalhados pelo citoplasma.

O aparelho de Golgi atua como centro de armazenamento, transformação, empacotamento e remessa de substâncias na célula. Muitas das substâncias que passam pelo aparelho de Golgi serão eliminadas da célula, indo atuar em diferentes partes do organismo. É o que ocorre, por exemplo, com asenzimas digestivas produzidas e eliminadas pelas células de diversos órgãos (estômago, intestino, pâncreas etc.). Outras substâncias, tais como o muco que lubrifica as superfícies internas do nosso corpo, também são processadas e eliminadas pelo aparelho de Golgi. Assim, o principal papel dessa estrutura citoplasmática é a eliminação de substâncias que atuam fora da célula, processo genericamente denominado secreção celular.
Secreção de enzimas digestivas
As enzimas digestivas do pâncreas, por exemplo, são produzidas no RER e levadas até as bolsas do aparelho de Golgi, onde são empacotadas em pequenas bolsas, que se desprendem dos dictiossomos e se acumulam em um dos pólos da célula pancreática. Quando chega o sinal de que há alimento para ser digerido, as bolsas cheias de enzimas se deslocam até a membrana plasmática, fundem-se com ela e eliminam seu conteúdo para o meio exterior.
A produção de enzimas digestivas pelo pâncreas é apenas um entre muitos exemplos do papel do aparelho de Golgi nos processos de secreção celular. Praticamente todas as células do corpo sintetizam e secretam uma grande variedade de proteínas que atuam fora delas.

O retículo endoplasmático
Tipos de retículo
O citoplasma das células eucariontes contém inúmeras bolsas e tubos cujas paredes têm uma organização semelhante à da membrana plasmática. Essas estruturas membranosas formam uma complexa rede de canais interligados, conhecida pelo nome de retículo endoplasmático. Pode-se distinguir dois tipos de retículo: rugoso (ou granular) e liso (ou agranular).
Retículo endoplasmático rugoso (RER) e liso (REL)
O retículo endoplasmático rugoso (RER), também chamado de ergastoplasma, é formado por sacos achatados, cujas membranas têm aspecto verrugoso devido à presença de grânulos – osribossomos – aderidos à sua superfície externa (voltada para o citosol). Já o retículo endoplasmático liso (REL) é formado por estruturas membranosas tubulares, sem ribossomos aderidos, e, portanto, de superfície lisa.

Tipos de retículo
O citoplasma das células eucariontes contém inúmeras bolsas e tubos cujas paredes têm uma organização semelhante à da membrana plasmática. Essas estruturas membranosas formam uma complexa rede de canais interligados, conhecida pelo nome de retículo endoplasmático. Pode-se distinguir dois tipos de retículo: rugoso (ou granular) e liso (ou agranular).
Retículo endoplasmático rugoso (RER) e liso (REL)
O retículo endoplasmático rugoso (RER), também chamado de ergastoplasma, é formado por sacos achatados, cujas membranas têm aspecto verrugoso devido à presença de grânulos – osribossomos – aderidos à sua superfície externa (voltada para o citosol). Já o retículo endoplasmático liso (REL) é formado por estruturas membranosas tubulares, sem ribossomos aderidos, e, portanto, de superfície lisa.
Os dois tipos de retículo estão interligados e a transição entre eles é gradual. Se observarmos o retículo endoplasmático partindo do retículo rugoso em direção ao liso, vemos as bolsas se tornarem menores e a quantidade de ribossomos aderidos diminuir progressivamente, até deixar de existir.
Funções do retículo endoplasmático
O retículo endoplasmático atua como uma rede de distribuição de substâncias no interior da célula. No líquido existente dentro de suas bolsas e tubos, diversos tipos de substâncias se deslocam sem se misturar com o citosol.
Produção de lipídios
Uma importante função de retículo endoplasmático liso é a produção de lipídios. A lecitina e o colesterol, por exemplo, os principais componentes lipídicos de todas as membranas celulares são produzidos no REL. Outros tipos de lipídios produzidos no retículo liso são os hormônios esteróides, entre os quais estão a testosterona e os estrógeno, hormônios sexuais produzidos nas células das gônadas de animais vertebrados.
Desintoxicação
O retículo endoplasmático liso também participa dos processos de desintoxicação do organismo. Nas células do fígado, o REL, absorve substâncias tóxicas, modificando-as ou destruindo-as, de modo a não causarem danos ao organismo. É a atuação do retículo das células hepáticas que permite eliminar parte do álcool, medicamentos e outras substâncias potencialmente nocivas que ingerimos.
Armazenamento de substâncias
Dentro das bolsas do retículo liso também pode haver armazenamento de substâncias. Os vacúolos das células vegetais, por exemplo, são bolsas membranosas derivadas do retículo que crescem pelo acúmulo de soluções aquosas ali armazenadas.
Produção de proteínas
O retículo endoplasmático rugoso, graças à presença dos ribossomos, é responsável por boa parte da produção de proteínas da célula. As proteínas fabricadas nos ribossomos do RER penetram nas bolsas e se deslocam em direção ao aparelho de Golgi, passando pelos estreitos e tortuosos canais co retículo endoplasmático liso.
Ribossomo
Função: síntese protéica.
Estrutura: existem dois tipos de ribossomos, um presente nos seres procariontes, cloroplastos e mitocôndrias e outro nos eucariontes. Os ribossomos são compostos por duas subunidades de tamanhos diferentes, que depois de prontas se separam e saem do núcleo pelo poro nuclear, passando para o citoplasma. Quando o ribossomo está disperso no citoplasma, recebe o nome de ribosoma livre e quando está aderido à superfície externa das estruturas membranosas, é chamado de ribosoma aderido.
Constituição: intensamente basófilos, compostos de quatro tipos de rRNA e 80 proteínas diferentes associadas, as quais unem-se para formar uma estrutura globular condensada.

Funções: digestão intracitoplasmática, renovação das organelas celulares, e metabolização de diversas moléculas. As substâncias do meio extracelular entram na célula através dos fagossomos. No interior da célula, o fagossomo se funde com o lisossomo primário. O processo de digestão inicia dentro de outro vacúolo, o lisossomo secundário.
Estrutura: vesículas delimitadas por membrana. Estão presentes em quase todas as células, mas em maior quantidade nos macrófagos. Geralmente são organelas esféricas e com aspecto granuloso.
Constituição: contém enzimas lisossômais (como: fosfatase ácida, glicuronidase, sulfatase, ribonuclease e colagenase) que variam com a célula. Estas enzimas são sintetizadas e segregadas no REG, transportas para o Aparelho de Golgi, onde são empacotadas, formando os lisossomos primários.Estrutura e origem dos lisossomos.
Os lisossomos (do grego lise, quebra, destruição) são bolsas membranosas que contêm enzimas capazes de digerir substâncias orgânicas. Com origem no aparelho de Golgi, os lisossomos estão presentes em praticamente todas as células eucariontes. As enzimas são produzidas no RER e migram para os dictiossomos, sendo identificadas e enviadas para uma região especial do aparelho de Golgi, onde são empacotadas e liberadas na forma de pequenas sas.
A digestão intracelular
Os lisossomos são organelas responsáveis pela digestão intracelular. As bolsas formadas na fagocitose e na pinocitose, que contêm partículas capturadas no meio externo, fundem-se aos lisossomos, dando origem a bolsas maiores, onde a digestão ocorrerá.
As bolsas originadas pela fusão de lisossomos com fagossomos ou pinossomos são denominadas vacúolos digestivos; em seu interior, as substâncias originalmente presentes nos fagossomos ou pinossomos são digeridas pelas enzimas lisossômicas.
À medida que a digestão intracelular vai ocorrendo, as partículas capturadas pelas células são quebradas em pequenas moléculas que atravessam a membrana do vacúolo digestivo, passando para o citosol. Essas moléculas serão utilizadas na fabricação de novas substâncias e no fornecimento de energia à célula.
Eventuais restos do processo digestivo, constituídos por material que não foi digerido, permanecem dentro do vacúolo, que passa a ser chamado vacúolo residual.
Muitas células eliminam o conteúdo do vacúolo residual para o meio exterior. Nesse processo, denominado clasmocitose, o vacúolo residual encosta na membrana plasmática e fundem-se com ela, lançando seu conteúdo para o meio externo.
Peroxissomos
Funções: oxidar substâncias orgânicas, prozudir peróxido de hidrogênio, participa da b-oxidação, exporta acetil-CoA para o citossol, participa da síntese de ácidos biliares e de colesterol.
Estrutura: esféricas, envolvidos por membrana. Ele se divide por fissão.
Constituição: enzimas como a catalase, a urato oxidase e a D-aminoácidos e não possuem DNA e RNA. A catalase é uma enzima muito importante, pois ela oxida substâncias tóxicas ao organismo, inclusive o álcool etílico.
Microtúbulos
Funções: oferecer rigidez na forma das células, manter os prolongamentos celulares, dar simetria à célula, servir de suporte para as células na locomoção, servir como base morfológica para centríolos, cílios, flagelos e corpúsculos basais.
Estrutura: formado por treze subunidades de a e b tubulina, chamadas de herodímero. Organizadas em forma de espiral e com comprimento e estabilidade variáveis. Possuem eventuais comunicações entre um microfilamento e outro. A parte central do microtúbulo é denominada de axonema.
Constituição: depende do local. Miosina no músculo estriado. No restante das células, em geral, são constituídos por filamentos finos de actina e filamentos grossos de miosina associados às organelas. Também outras proteínas são visualizadas nos filamentos intermediários, que são constituídos por queratinas, por vimentina, por desmina, por proteína fibrilar ácida da glia e por proteínas dos neurofilamentos.
Muitos cientistas estão convencidos de que as mitocôndrias são descendentes de seres procariontes primitivos que um dia se instalaram no citoplasma das primeiras células eucariontes. Existem evidências que apóiam essa hipótese, tais como o fato de as mitocôndrias apresentarem material genético mais parecido com a das bactérias do que com a das células eucariontes em que se encontram
O mesmo ocorre com relação a maquinaria para a síntese de proteínas: os ribossomos mitocondriais são muito semelhantes aos das bactérias e bem diferentes dos ribossomos presentes no citoplasma das células eucariontes.
Centríolos
Os centríolos são organelas NÃO envolvidas por membrana e que participam do progresso de divisão celular. Nas células de fungos complexos, plantas superiores (gimnospermas e angiospermas) e nematóides não existem centríolos. Eles estão presentes na maioria das células de animas, algas e vegetais inferiores como as briófitas (musgos) e pteridófitas (samambaias).
Estruturalmente, são constituídos por um total de nove trios de microtúbulos protéicos, que se organizam em cilindro.
São autoduplicáveis no período que precede a divisão celular, migrando, logo a seguir, para os pólos opostos da célula.
Uma das providências que a fábrica celular precisa tomar é a construção de novas fábricas, isto é, a sua multiplicação. Isso envolve uma elaboração prévia de uma serie de “andaimes” protéicos, o chamado fuso de divisão, formado por inúmeros filamentos de microtúbulos.
Embora esses microtúbulos não sejam originados dos centríolos e sim de uma região da célula conhecido como centrossomo, é comum a participação deles no processo de divisão de uma célula animal. Já em células de vegetais superiores, como não existem centríolos, sua multiplicação se processa sem eles.

Os Cílios e Flagelos
São estruturas móveis, encontradas externamente em células de diversos seres vivos. Os cílios são curtos e podem ser relacionados à locomoção e a remoção de impurezas. Nas células que revestem a traquéia humana, por exemplo, os batimentos ciliares empurram impurezas provenientes do ar inspirado, trabalho facilitado pela mistura com o muco que, produzido pelas células da traquéia, lubrifica e protege a traquéia. Em alguns protozoários, por exemplo, o paramécio, os cílios são utilizados para a locomoção
Os flagelos são longos e também se relacionam a locomoção de certas células, como a de alguns protozoários (por exemplo, o tripanosssomo causador da doença de Chagas) e a do espermatozóide.

Em alguns organismos pluricelulares, por exemplo, nas esponjas, o batimento flagelar cria correntes de água que percorrem canais e cavidades internas, trazendo, por exemplo, partículas de alimento.
Estruturalmente, cílios e flagelos são idênticos. Ambos são cilíndricos, exteriores as células e cobertos por membrana plasmática. Internamente, cada cílio ou flagelo é constituído por um conjunto de nove pares de microtúbulos periféricos de tubulina, circundando um par de microtúbulos centrais. É a chamada estrutura 9 + 2.

Tanto os cílios como flagelos são originados por uma região organizadora no interior da célula, conhecida como corpúsculo basal. Em cada corpúsculo basal há um conjunto de nove trios de microtúbulos (ao invés de duplas, como nos cílios e flagelos), dispostos em círculo. Nesse sentido, a estrutura do corpúsculo basal é semelhante à de um centríolo.
Citoesqueleto
Quando se diz que o hialoplasma é um fluido viscoso, fica-se com a impressão de que a célula animal tem uma consistência amolecida e que se deforma a todo o momento. Não é assim.
Um verdadeiro “esqueleto” formado por vários tipos de fibras de proteínas cruza a célula em diversas direções, dando-lhe consistência e firmeza.
Essa “armação” é importante se lembrarmos que a célula animal é desprovida de uma membrana rígida, como acontece com a membrana celulósica dos vegetais.
Entre as fibras protéicas componentes desse “citoesqueleto” podem ser citados os microfilamentos de actina, os microtúbulos e os filamentos intermediários.

Os microfilamentos são os mais abundantes, constituídos da proteína contráctil actina e encontrados em todas as células eucarióticas. São extremamente finos e flexíveis, chegando a ter 3 a 6 nm (nanômetros) de diâmetro, cruzando a célula em diferentes direções , embora concentram-se em maior número na periferia, logo abaixo da membrana plasmática. Muitos movimentos executados por células animais e vegetais são possíveis graças aos microfilamentos de actina.
 |
| Add caption |
Os microtúbulos, por sua vez, são filamentos mais grossos, de cerca de 20 a 25 nm de diâmetro, quefuncionam como verdadeiros andaimes de todas as células eucarióticas. São, como o nome diz, tubulares, rígidos e constituídos por moléculas de proteínas conhecidas como tubulinas, dispostas helicoidalmente, formando um cilindro. Um exemplo, desse tipo de filamento é o que organiza o chamado fuso de divisão celular. Nesse caso, inúmeros microtúbulos se originam e irradiam a partir de uma região da célula conhecida como centrossomo (ou centro celular) e desempenham papel extremamente importante na movimentação dos cromossomos durante a divisão de uma célula.

Outro papel atribuído aos microtúbulos é o de servir como verdadeiras “esteiras” rolantes que permitem o deslocamento de substâncias, de vesículas e de organóides como as mitocôndrias e cloroplastos pelo interior da célula. Isso é possível a partir da associação de proteínas motoras com os microtúbulos.
Essas proteínas motoras ligam-se de um lado, aos microtúbulos e, do outro, à substância ou organóide que será transportado, promovendo o seu deslocamento.
Por exemplo, ao longo do axônio (prolongamento) de um neurônio, as proteínas motoras conduzem, ao longo da “esteira” formada pelos microtúbulos, diversas substâncias para as terminações do axônio e que terão importante participação no funcionamento da célula nervosa.

Filamentos intermediários
Os filamentos intermediários são assim chamados por terem um diâmetro intermediário – cerca de 10 nm – em relação aos outros dois tipos de filamentos protéicos.

Nas células que revestem a camada mais externa da pele existe grande quantidade de um tipo de filamento intermediário chamado queratina. Um dos papeis desse filamento é impedir que as células desse tecido se separem ou rompam ao serem submetidas, por exemplo, a um estiramento.
Além de estarem espalhadas pelo interior das células, armando-as, moléculas de queratina promovem uma “amarração” entre elas em determinados pontos, o que garante a estabilidade do tecido no caso da ação de algum agente externo que tente separá-las. Esse papel é parecido ao das barras de ferro que são utilizadas na construção de uma coluna de concreto. Outras células possuem apreciável quantidade de outros filamentos intermediários. É o caso das componentes dos tecidos conjuntivos e dos neurofilamentos encontrados no interior das células nervosas.
RESUMO:

Citologia. Disponível em:http://www.sobiologia.com.br/conteudos/Citologia/cito.php. Acessado em 20 de março de 2012.
FONSECA, Krukemberghe.Núcleo das células. Disponível em:http://www.brasilescola.com/biologia/nucleo-das-celulas.htm . Acessado em 20 de março de 2012.
Assinar:
Postagens (Atom)